论文标题:Harnessing accurate non-homologous end joining for efficient precise deletion in CRISPR/Cas9-mediated genome editing
期刊:Genome Biology
作者:Tao Guo, Yi-Li Feng, Jing-Jing Xiao, Qian Liu, Xiu-Na Sun,Ji-Feng Xiang, Na Kong, Si-Cheng Liu, Guo-Qiao Chen, Yue Wang, Meng-Meng Dong, Zhen Cai, Hui Lin, Xiu-Jun Cai and An-Yong Xie
发表时间:2018/10/19
数字识别码:10.1186/s13059-018-1518-x
原文链接:https://genomebiology.biomedcentral.com/articles/10.1186/s13059-018-1518-x?utm_source=WeChat&utm_medium=Website_linksSocial_media_organic&utm_content=CelZha-MixedBrand-multijournal-Multidisciplinary-China&utm_campaign=ORG_AWA_CZH_BMCWechat_dailyposts_blogs
微信链接:https://mp.weixin.qq.com/s/r5wl6JdZ9xaPVKWt99Nn1w
CRISPR基因编辑技术主要是通过细胞内DNA双链断裂(Double strand breaks,DSB)的定点诱导及随后的修复来实现,其中可利用的一条主要修复途径是非同源末端连接(non-homologous end joining,NHEJ)。由于CRISPR/Cas9介导的常规基因敲除效率高,且依赖于突变型NHEJ,修复CRISPR/Cas9诱导的DSB的NHEJ常被认为是易错的。然而,Cas9诱导的DSB主要是平末端,不需要额外的末端加工就可以直接相连,浙江大学医学院附属邵逸夫医院及转化医学研究院谢安勇研究组推测,CRISPR/Cas9诱导的NHEJ修复应该是偏向精准型的,而且这个偏向性或许可以帮助提高CRISPR/Cas9基因编辑的效率及精准度。尽管作者先前的研究已显示,可直接相连的DNA末端,包括Cas9 诱导产生的平末端,可以通过精准型NHEJ高效修复[1,2],但研究只局限于个别基因组位点,还未在多个位点上进行系统验证及分析,其中的规律也不明了。
为解决这个问题,该研究组设计了配对sgRNA靶向小鼠和人基因组的71个位点,引导Cas9在这些靶点上诱导两个邻近的DSB。这两个临近的DSB可以同时或分别产生:同时产生的DSB归类于Group I,只产生第一个DSB的为Group II,只产生第二个DSB的为Group III,两个DSB分别产生的为Group IV(图1a)。这些类型的DSB被NHEJ修复之后,通过靶点序列PCR扩增子深度测序,结合研究组开发的生信分析软件,可以计算不同类型修复产物频率,鉴定修复特征。在Group I DSB的修复中,无论NHEJ是精准型还是突变型,都将导致间隔序列丢失,因此通过分析接口序列可以将精准型NHEJ从突变型中区分开来。 结果显示,70个基因组位点总基因编辑效率(即NHEJ修复频率)平均约为35.8%,其中大约60%为Group I NHEJ产物,而Group I NHEJ产物中精准型NHEJ比例平均约50%(图1b),并且在很多位点上精准型NHEJ甚至超过80%。以上结果表明,修复Cas9产生的DSB的NHEJ途径偏向精准型NHEJ。正因为配对Cas9-sgRNA策略可以区分精准型NHEJ和突变型NHEJ,该方法还可以取代常用的NHEJ报告系统,用于定量分析细胞内和动物体内的NHEJ,而且更方便、更灵活。
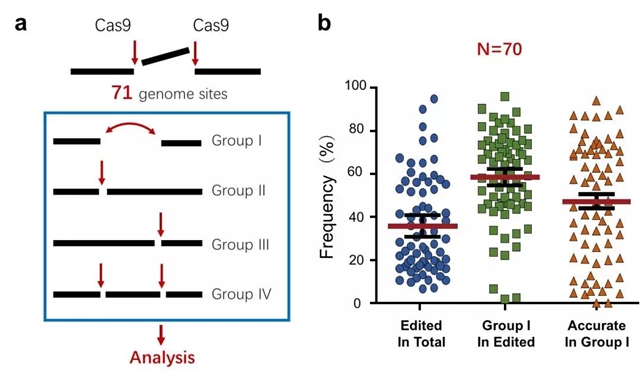
图1. 配对Cas9-sgRNA技术在71个内源基因组位点的NHEJ分析。a. 配对Cas9-sgRNA诱导的NHEJ产物(基因组编辑)示意图。b. 70个基因组位点的总编辑效率、Group I NHEJ效率及精准型NHEJ效率。额外的1个位点只能计算精准型NHEJ效率而未被包括在内。
课题组还注意到,在所分析的70个位点中,有的位点精准型NHEJ频率极低,甚至不超过20%(图1b),这样的现象通常被认为是受末端加工导致核苷酸丢失(即删除)影响所致。然而,分析显示,在修复Cas9诱导的DSB时,低频精准型NHEJ主要伴随的是高频核苷酸插入,而且这些核苷酸不是随机的,而基本都是由模板序列决定的核苷酸插入,称模板化插入(templated insertions,TI)(图2a)。先前的研究已表明,Cas9的RuvC和HNH核酸酶结构域通常在PAM(protospacer adjacent motif)前第3位碱基切割,产生平末端,但RuvC有时在第4位或第5位碱基切割,产生带1个 或2 个核苷酸(nucleotides,nt)外伸端的5’粘性末端[3]。此时,由于Group I DSB的两个5’粘性末端有极小的互补概率,细胞难以通过精准型NHEJ修复,而是通过DNA聚合酶将粘性末端补平,在连接时就会产生+1核苷酸或+2核苷酸的模板化插入[4]。这些模板化插入的频率与精准型NHEJ发生频率成负相关,即精准型NHEJ的效率越低,模板化插入比例越高(图2b)。也因为这个原因,配对Cas9-sgRNA策略所得出的精准型NHEJ频率低估了修复Cas9诱导的单个DSB的精准型NHEJ效率。
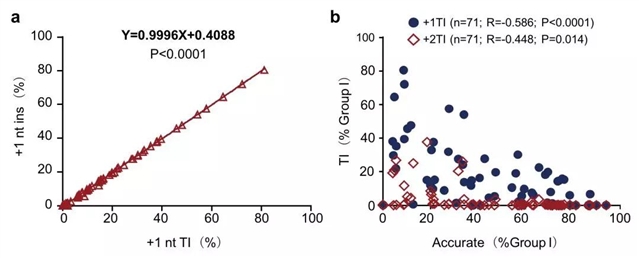
图2. 71个基因组位点模板化插入对精准型NHEJ效率的影响。a. +1 nt插入效率与+1 nt模板化插入效率几乎完全正相关。b. +1 nt和+2 nt模板化插入效率与精准型NHEJ效率呈负相关。
那么,是否可以通过避免模板化插入提高精准型NHEJ的效率呢?根据模板化插入产生的原理,配对Cas9-sgRNA在靶点上的特定定位应该可以避免模板化插入。Cas9-sgRNA在靶点上的定位由PAM决定,PAM 基序既可以在Watson链(即W)上,也可以在Crick链(即C)上。因此,配对Cas9-sgRNA在靶点上的位置可以排列出四种组合:W/W、W/C、C/W和C/C(图3a)。如果Cas9产生的DSB是平末端,修复产物不会有高频的模板化插入;只有当Cas9产生5’外伸端时通过DNA聚合酶补平才会产生模板化插入。然而,在W/C位置组合中,前后两个DNA末端的5’外伸端补平后相当于Cas9切割产生的平末端,可以预测修复完成后不会有额外的模板化插入,而其它三种组合(W/W、C/W和C/C)则会产生(图3a)。结果正如预期,通过对71个基因组位点的分析,配对Cas9-sgRNA为W/C位置时,几乎检测不到模板化插入(图3b),而且精准型NHEJ效率最高(图3c)。这一结果提示在使用配对Cas9-sgRNA基因编辑技术时,尽可能选择“W/C”位置组合以期获得最高效率的精准型NHEJ修复事件。
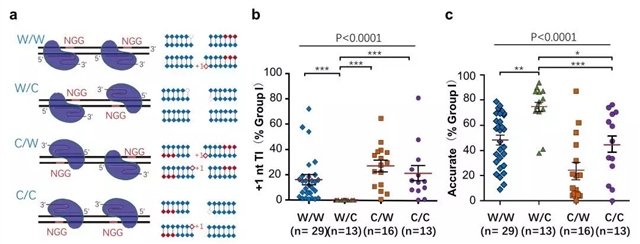
图3. 配对Cas9-sgRNA的PAM组合对精准型NHEJ的影响。a. 根据配对Cas9-sgRNA在靶点上的PAM位置产生的四种组合。b. 配对Cas9-sgRNA的PAM组合对模板化插入产生的影响。W/C组合几乎不产生模板化插入。c. 配对Cas9-sgRNA的四种PAM组合对精准型NHEJ效率的影响。W/C组合产生更高效的精准型NHEJ。